DINOSAURICON G
september 30, 2012 1 reactie
G
- “Gadolosaurus” – nomen nudum
- Galesaurus – actually a therapsid
- Gallimimus Gallimimus
http://library.thinkquest.org/C0128701/Pictures/gallimimus.jpg http://www.nhm.ac.uk/nature-online/life/dinosaurs-other-extinct-creatures/ostrich-dino-article/index.html
Gallimimus June 15, 2011
Filed under: Theropoda —
With a short, light body and long back legs, Gallimimus was a fast-running dinosaur. It took very long strides and could outrun most predators. It looked like a large ostrich with its long neck and toothless beak. Its stiff tail helped it to balance when running.

Factbox//Name: Gallimimus, meaning ‘chicken mimic’ Size: 6m long and 3m high Food: plants, eggs, insects and lizards Lived: about 70 million years ago in the Cretaceous Period in Mongolia

Gallimimusis the largest known type of ornithomimid, but it has shorter arms in proportion to the other species. The hands, too, are quite small and the fingers are not very flexible. The head is quite long and graceful and, as in nearly all ornithomimids, the jaws have no teeth. The beak of the lower jaw is shovel-shaped, and the big eyes are situated on the sides of the head, so it did not have binocular vision.Like the other ostrich mimics and modern birds, Gallimimus had hollow bones. This device allowed for a reduction of weight in the body, without reducing the strength, and enabled the animal to move quickly. The main difference between the two known Gallimimus species is the shape of the fingers. G. mongoliensishad shorter hands and would not have grasped as well.Gallimimus had short arms with three claws on its hands. The claws were sharp, but Gallimimuscould not grasp things very well and did not eat meat because it could not tear it up.Gallimimus’claws came in very useful, however, because it used them to scrape away at the soil to dig up eggs for food. It ate mostly plants, but it also fed on small insects, which it grabbed in its beak, and even chased lizards.

- Galtonia – actually a pseudosuchian; possibly a junior synonym of Revueltosaurus

http://scienceblogs.com/tetrapodzoology/2007/05/24/galve-european-spinosaurines-c/
- Galvesaurus – junior synonym of Galveosaurus
- “Gansutitan” – nomen nudum; Daxiatitan
Ornithischia Thyreophora Ankylosauria Ankylosauridae?
Subfamily: Polacanthinae?
Genus: Gargoyleosaurus
Species: G. parkpinorum
Fossil range: Late Jurassic

Gargoyleosaurus (meaning “gargoyle lizard”) is one of the earliest ankylosaurs known from reasonably complete fossil remains. Its skull measures 29 centimetres (11 in) in length, and its total body length is an estimated 3 to 4 metres (9.8 to 13 ft). It may have weighed as much as 1 tonne (2,200 lb). The holotype was discovered at the Bone Cabin Quarry West locality, in Albany County, Wyoming in exposures of the Upper Jurassic (Kimmeridgian to Tithonian stages) Morrison Formation.
The type species, G. parkpinorum (originally G. parkpini) was described by Ken Carpenter et al. in 1998. A mounted skeletal reconstruction of Gargoyleosaurus parkpinorum can be seen at the Denver Museum of Nature and Science. Gargoyleosaurus was present in stratigraphic zone 2 of the Morrison Formation.

Holotype
The holotype specimen of Gargoyleosaurus parkpinorum was collected by Western Paleontology Labs in 1996 and is currently held in the collections of the Denver Museum of Nature and Science, Denver, Colorado. Besides the holotype, two other partial skeletons are known (although not yet described) The holotype consists of most of the skull and a partial postcranial skeleton. The specimen was originally described as Gargoyleosaurus parkpini by Carpenter, Miles and Cloward in 1998, then renamed G. parkpinorum by Carpenter et al. in 2001, in accordance with ICZN art. 31.1.2A.

Classification
Much of the skull and skeleton has been recovered, and the taxon displays cranial sculpturing, including pronounced deltoid quadratojugal and squamosal bosses. The taxon is further characterized by a narrow rostrum (in dorsal view), the presence of seven conical teeth in each premaxilla, an incomplete osseous nasal septum, a linerarly arranged nasal cavity, the absence of an osseus secondary palate, and, as regards osteoderms, two sets of co-ossified cervical plates and a number of elongate conical spines.
Vickaryous et al. (2004) place Gargoyleosaurus parkpinorum within the Family Ankylosauridae of the Ankylosauria and are in agreement with most previous phylogenetic hypotheses, which place the genus as the sister group to all other ankylosaurids (i.e., members of the Ankylosauridae). These studies however, only utilized the skull, whereas many of the distinctive features of the family Polacanthidae are in the postcranial skeleton.
http://carnivoraforum.com/topic/9329337/1/

This great photo of the holotype of Gargoyleosaurus parkpini, DMNH 27726, was taken by Jim Puckett in the Dinosaur Hall of “Prehistoric Journey” at the Denver Museum of Nature and Science.
 Reconstructed skeleton of the Jurassic ankylosaur (Gargoyleosaurus parkpini) in the Denver Museum of Science and Nature. Note scutes dermal covering the head and upper body.)
Reconstructed skeleton of the Jurassic ankylosaur (Gargoyleosaurus parkpini) in the Denver Museum of Science and Nature. Note scutes dermal covering the head and upper body.)
- GarudimimusGarudimimus is een geslacht van theropode dinosauriërs, behorend tot de groep van de Maniraptoriformes, dat tijdens het late Krijt leefde in het gebied van het huidige Mongolië. De enige benoemde soort is Garudimimus brevipes

Garudimimus brevipes Barsbold, 1981
Garudimimidé du Cenomanien/Santonien (99.6-83.5 MA) de Mongolie (Ömnögov’); identifié par un squelette partiel (crâne, os des bras, vertèbres, pubis …).


Garudimimus, meaning “Garuda mimic”, is a genus of ornithomimosaur dinosaur from the Upper Cretaceous Period. It was found in sedimentary deposits in Bayshin Tsav, Mongolia.
Garudimimus was about 13 feet in length and was possibly an omnivore. It is believed that this dinosaur was not built for speed, unlike most ornithomimosaurs. It had short legs and heavy feet. The muscles of the legs were not as developed as in more derived ornithomimosaurs. Each foot had four toes, also unlike the typical three-toed relatives. The skull was rounded and the eyes would have been large. It was previously thought that this dinosaur had a horn at the top of the skull, however, recent studies show that the horn was a misplaced skull bone.

http://www.dinosaurus.net/genera/GGG/gasosaurus.htm
http://www.nhm.ac.uk/nature-online/life/dinosaurs-other-extinct-creatures/dino-directory/detail.dsml?Genus=Gasosaurus
http://www.dinodata.info/index.php?topic=1487.0
- Gasparinisaura Gasparinisaura is een geslacht van plantenetende dinosauriërs, behorend tot de groep van de Euornithopoda, dat tijdens het late Krijt leefde in het gebied van het huidige Zuid-Amerika. De enige benoemde soort is Gasparinisaura cincosaltensis.


- Gastonia Gastonia Gastonia is een uitgestorven geslacht van plantenetende ornithischische dinosauriërs dat tijdens het Vroeg-Krijt leefde in het gebied van het huidige Noord-Amerika.



http://blog.press.princeton.edu/2010/10/02/pgs-daily-dinosaur-gastonia-burgei/
| A few skulls and skeletons from nearly complete to partial. | |
| ANATOMICAL CHARACTERISTICS | Head very small, heavily armored, no teeth on front of upper jaw. Arm and leg very short. Belly extremely broad. Large sidewaysprojecting shoulder spines, no lateral spines at hip, modest spines on side of tail. |
| AGE | Early Cretaceous, Barremian. |
| DISTRIBUTION AND FORMATION | Utah; Lower Cedar Mountain. |
| HABITAT | Short wet season, otherwise semiarid with floodplain prairies, open woodlands, and riverine forests |
- “Gavinosaurus” – nomen nudum; Eotyrannus
- Geminiraptor
- Genusaurus
- Genyodectes
- Geranosaurus
____________________________________________________________________________________________________
- Giganotosaurus Giganotosaurus.
Giganotosaurus - http://www.tunturisusi.com/dinosaurukset/giganotosaurus.jpg

http://www.dinosaurus.net/genera/GGG/giganotosaurus.htm
http://www.dinosaur-facts.com/on-the-ground/giganotosaurus/


-
FOSSILMUSEUMThe Giganotosaurus from Patagonia,was 10% bigger than T’ Rex
http://www.fossilmuseum.net/DinosaurFossils/giganotosaurus/giganotosaurus.jpg _______ Giganotosaurus carolinii

-
Gigantosaurus carolinii, bigger than the Mapusaurus , it was even larger than the Tyranosaurus Rex, with a skull 1.8m long.________________________________________________________________________________________________
- Gigantoraptor

Gigantoraptor Erlianensis
- Gigantosaurus Seeley, 1869
- “Gigantosaurus” E. Fraas, 1908 – preoccupied name, now known as Tornieria, Malawisaurus, and Janenschia
- Gigantoscelus – junior synonym of Euskelosaurus
- Gigantspinosaurus Gigantspinosaurus is een geslacht van plantenetende ornithischische dinosauriërs, behorend tot de groep van de Stegosauria, dat tijdens het Opper-Jura leefde in het gebied van het huidige China


- Gilmoreosaurus
- “Ginnareemimus” – nomen nudum; Kinnareemimus


Berlin: MFN Dinosaur Hall,
It and the other Brachiosaurids are members of the Brachiosauridae family of dinosaurs.They differ from other sauropods, as all had a long giraffe-like build, with long forelimbs and a raised neck, which it probably used to graze in the tops of trees. It has been suggested that all were basal titanosauriforms thrown together without respect for true characteristics, such as long necks and long arms.
For many decades, Brachiosaurus was the largest known dinosaur. It has since been exceeded in sheer mass by a number of giant titanosaurids like the Argentinosaurus and it was finally surpassed in height by another brachiosaurid, the Sauroposeidon. It was, however, still the largest dinosaur known from a relatively complete skeleton and the largest on display anywhere in the world.
The first Brachiosaurus was discovered in 1900 by Elmer S. Riggs, in the Grand River Canyon of western Colorado, in the United States. He named the new species and genera in 1903 after its long front limbs. Brachiosaurus means “arm lizard”, from the Greek brachion (“arm”) and sauros (“lizard”). This first discovered species has named Brachiosaurus altithorax and it was made famous by the movie “Jurassic Park”. It is very tall (40 feet!) and very heavy – more than 12 elephants! Unlike most of the other long-necked dinosaurs,
Starting in 1909, German paleontologist Werner Janensch found many new specimens in Tanzania, Africa, including some nearly complete skeletons. These were the Brachiosaurus brancai, which is shown on the poster. Its fossilized remains are on display at the Humboldt Museum in Berlin, Germany. The remains are primarily from one gigantic animal, except for a few tail bones (caudal vertebrae) which belong to another animal of the same size and species While the Diplodocus carnegiei mounted at the Carnegie Museum of Natural History in Pittsburgh, United States actually exceeds it in length, the Berlin animal is taller, and far more massive. Almost a century after its discovery, it still remains the largest mounted dinosaur in the world. It has also been one of the luckiest, because it escape destruction when most of Berlin was reduced to rubble by allied bombardment during World War II.There are three known species of Brachiosaurus;
B. alataiensis (de Lapparent & Zbyszewski, 1957): Is known from back bones (vertebrae), and parts of the hip and limbs, which were recovered in Estremadura, Portugal. It lived about 150 million years ago, during the Kimmeridgian age of the late Jurassic period.
B. altithorax (Riggs, 1903). The type species is known from two partial skeletons recovered in Colorado and Utah in the United States. It lived from 145 to 150 million years ago, during the Kimmeridgian to Tithonian ages.
?B. nougaredi (de Lapparent, 1960): While it may not be a distinct species (nomen dubium?) it is known from set of fused bones over the hip (sacrum), and parts of a forelimb, which were recovered in Wargla, Algeria in Africa. It lived 100 to 110 million years ago, during the Albian to Cenomanian ages of the middle Cretaceous period.There at least two other valid members of the family. :
Giraffatitan brancai (Janensch, 1914 (formerly B. brancai): The new type species, it is known from five partial skeletons, including at least three skulls, and some limb bones, which were recovered in Mtwara, Tanzania, in Africa. It lived from 145 to 150 million years ago, during the Kimmeridgian to Tithonian ages of the late Jurassic period.
Cedarosaurus weiskopfae (Tidwell, Carpenter, and Brooks, 1999). A new brachiosaurid from the Yellow Cat member of the Cedar Mountain Formation in Utah. This sauropod is known from a partial skeleton including vertebrae, partial girdles, and most of the limbs. Remains referred to Pleurocoelus may belong here. It was a smallish sauropod.http://www.dinosaur-world.com/weird_dinosaurs/giraffatitan%20_brancai.htm

This is (tentatively) identified as a plant-eating, longnecked dinosaur, or sauropodomorph.
Again, this was a big eater.
Illustration by Mike Belknap

Skeleton of Giraffatitan.
- Gobiceratops
- Gobipteryx – actually an enantiornithine bird
- Gobisaurus
- Gobititan
- “Godzillasaurus” – nomen nudum; Gojirasaurus
- Gojirasaurus
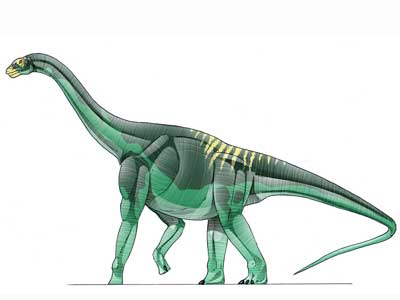
Gondwanatitan is a Primitive Titanosaur from late Cretaceous Brazil. It was similar to Aeolosaurus.

http://dinossaurosondeviveramcaracteristicas.blogspot.be/2013_06_01_archive.html
http://dinosaurs.about.com/od/herbivorousdinosaurs/p/gondwanatitan.htm
- Gongbusaurus Gongbusaurus is een geslacht van plantenetende ornithischische dinosauriërs dat tijdens het late Jura leefde in het gebied van het huidige China


“Gongbusaurus” wucaiwanensis
Gongbusaurus (meaning “Yu Gong’s (of the Zigong Dinosaur Museum) lizard”) is a genus of ornithischian (an ornithopod?) dinosaur that lived between about 160 and 155 million years ago, in the Late Jurassic period. A small herbivore, it is very poorly known. Two species have been assigned to it, but as the original name is based on teeth, there is no concrete evidence to connect the two species. Its fossils have been found in China.

Afbeeldingen van Gongxianosaurus

Skeletal reconstruction of Gongxianosaurus shibeiensis. From He et al., 1998.
http://www.palaeocritti.com/by-group/dinosauria/sauropoda/gongxianosaurus
 Gorgosaurus libratus
Gorgosaurus libratusGorgosaurus, meaning “fierce lizard” (from the Greek: gorgos/ meaning ‘terrible’ or ‘fierce’ and saurus/ meaning ‘lizard’) is a genus of carnivorous dinosaur that reached 7 to 8 metres (27 to 30 feet) in length, with an estimated weight of 2.5 tonnes (2.75 short tons). It was first described by paleontologist Lawrence Morris Lambe, in 1914 and has been found in western Canada and the United States. It lived about 70 million years ago in the late Cretaceous Period.
Over 20 Gorgosaurus skeletons have been recovered, making it the most well-represented tyrannosaurid in the fossil record. Generally similar to Tyrannosaurus and most other large tyrannosaurids (such as Daspletosaurus and Albertosaurus), Gorgosaurus can be described as having a massive head, large curved teeth, tiny two-fingered front limbs, and powerful legs. Compared to the other tyrannosaurids, Gorgosaurus is most similar to its very close relative Albertosaurus. Although it has been suggested that Gorgosaurus was a scavenger, its co-existence with the similarly sized but more robust tyrannosaurid, Daspletosaurus, casts doubt on this theory. Another hypothesis proposes that Gorgosaurus, which was rather lean for a tyrannosaurid, actively hunted fleet-footed animals such as duckbills and ornithomimids (‘ostrich-mimic’ dinosaurs). According to this proposition, the more troublesome ceratopsians and ankylosaurians (horned and heavily armoured dinosaurs) would have been left to Daspletosaurus.For years, the species Gorgosaurus libratus (the only species of Gorgosaurus currently recognized) was assigned to the Albertosaurus genus. However, recent work done by paleontologists suggest that enough differences exist between G. libratus and the other Albertosaurus species, to justify the original genus name of Gorgosaurus.Order: Saurischia
Suborder: Theropoda
Family: Tyrannosauridae
Subfamily: Albertosaurinae
Genus: Gorgosaurus
Species: G. libratus
 Image © The Natural History Museum, London 2007
Image © The Natural History Museum, London 2007
Goyocephale was a genus of dinosaur which lived during the Late Cretaceous period. It lived in what is now Mongolia. Goyocephale was a pachycephalosaur, which were ornithischians with thick, bony skulls.Goyocephale probably weighed 10-40 kg. The type species, Goyocephale lattimorei, was formally described by Perle, Maryanska, and Osmolska in 1982. It is based on a fairly complete skeleton.
Order: Ornithischia
Suborder: Marginocephalia
Infraorder: Pachycephalosauria
Family: Pachycephalosauridae
Genus: Goyocephale
Perle, Maryanska, and Osmolska (1982)
Species:
Goyocephale lattimorei
- Graciliceratops
- Graciliraptor
- Gracilisuchus – actually a non-dinosaurian archosaur
- “Gravisaurus” – nomen nudum; Lurdusaurus
- Gravitholus
- Gresslyosaurus – junior synonym of Plateosaurus
- Griphornis – junior synonym of the bird Archaeopteryx
- Griphosaurus – junior synonym of the bird Archaeopteryx
- “Gripposaurus” – nomen nudum; “Gyposaurus” sinensis
- Gryphoceratops
- “Gryphognathus” – nomen nudum; Gryphoceratops


Gryponyx africanus
Gryponyx taylori
Gryponyx transvaalensis
A new species of the hadrosaurine hadrosaurid Gryposaurus was discovered in the late Campanian Kaiparowits Formation of southern Utah.
Gryposaurus monumentensis
Lees meer … Palaeoblog
Lees het oorspronkelijke wetenschappelijke artikel
http://onlinelibrary.wiley.com/doi/10.1111/j.1096-3642.2007.00349.x/full


Gryposaurus notabilis
Gryposaurus was a common duck-billed dinosaur known from a over 10 skulls, some bones, and one of the very few skin casts ever found. The skin cast reveals the skin on its neck, sides and belly were covered with smooth scales less than a quarter of an inch in size. It had a long, narrow skull, highly-arched nostrils, and a big bump on its snout. It was a large plant-eater that would have traveled in herds while trying to avoid being eaten by some of the earliest tyrannosaur family members. Discovered in Alberta, Canada by L. Lambe in 1914, the type species is G. notabilis.
Gryposaurus is much better known under the name Kritosaurus notabilis. For a long time Kritosaurus and Gryposaurus were considered to be the same genus, but in the last twenty years it has become clear that the type material of Kritosaurus is not particularly diagnostic, based on a fragmentary skull.
Gryposaurus, meanwhile, is based on much better remains. Interestingly, it now appears that Kritosaurus may actually be a “saurolophinid”, instead of a “gryposaurinid”, illustrating the problems of establishing hadrosaurid taxa on material that is either missing or has an inadequately preserved skull.
hadrosaurid, hadrosaur, duck-billed dinosaur= was a plant-eater that was about 30 feet (9 m) long.
- Guaibasaurus Guaibasaurus is een geslacht van saurischische dinosauriërs dat tijdens het late Trias leefde in het gebied van het huidige Brazilië. De typesoort is Guaibasaurus candelariensis.
http://maniraptora.com/dinosaurs_world/macn/guaiba/code.html

Guaibasaurus (“Rio Guaiba lizard”) was an early dinosaur from the Late Triassic, about 220 million years ago. It was found in 1998 by José Bonaparte and Jorge Ferigolo near Candalaria, Río Grande do Sul Province, Brazil. The remains consist of two incomplete skeletons. Guaibasaurus is an even more primitive saurischian than either the Argentinean Herrerasaurus or the Brazilian Staurikosaurus, and may belong to the group that was ancestral to both sauropods and theropods.

- Guanlong .
.
.
.
.
- Gwyneddosaurus – a tanystrophid
- Gyposaurus – junior synonym of Massospondylus
http://en.wikipedia.org/wiki/List_of_dinosaurs




.jpg)








 Discovered in England and described by Sir Richard Owen in 1861, Echinodon was once considered to be a fabrosaurid, a family name that has now been abandoned. It may have been related to Scutellosaurus, a plant-eater with small bony armor plates on its back. Evidence suggests that armor found in England in 1879 and thought to belong to a lizard, may belong to Echinodon, thus moving it into the mostly quadrupedal Thyreophora suborder of ornithischians.
Discovered in England and described by Sir Richard Owen in 1861, Echinodon was once considered to be a fabrosaurid, a family name that has now been abandoned. It may have been related to Scutellosaurus, a plant-eater with small bony armor plates on its back. Evidence suggests that armor found in England in 1879 and thought to belong to a lizard, may belong to Echinodon, thus moving it into the mostly quadrupedal Thyreophora suborder of ornithischians.














 Skin impression from the abdomen of Edmontosaurus annectens
Skin impression from the abdomen of Edmontosaurus annectens










 Interactions with theropods
Interactions with theropods

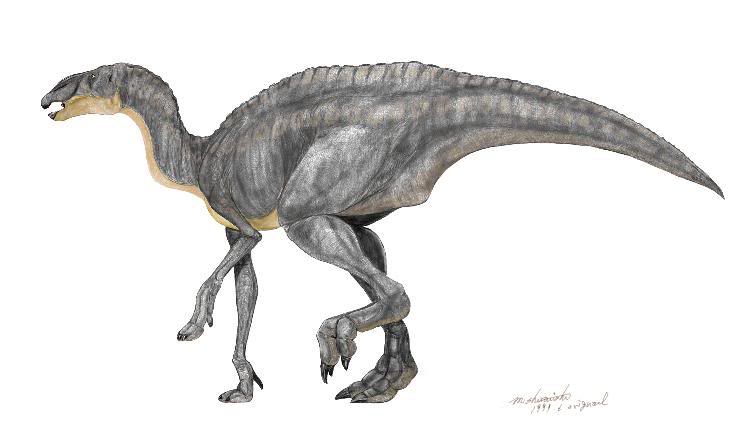




 Einiosaurus had a large downward-curving nasal horn.
Einiosaurus had a large downward-curving nasal horn.


















 Life restoration of
Life restoration of 



")




















")


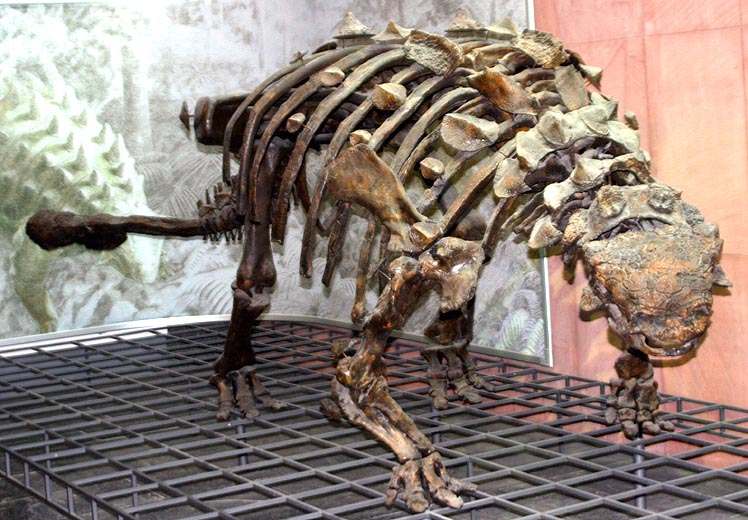
 this rather lovely centrosaurus skeleton. nearly complete from head to toe… it had toe bones. this skeleton apart from some vertebrae was complete
this rather lovely centrosaurus skeleton. nearly complete from head to toe… it had toe bones. this skeleton apart from some vertebrae was complete
 torosaurus
torosaurus






































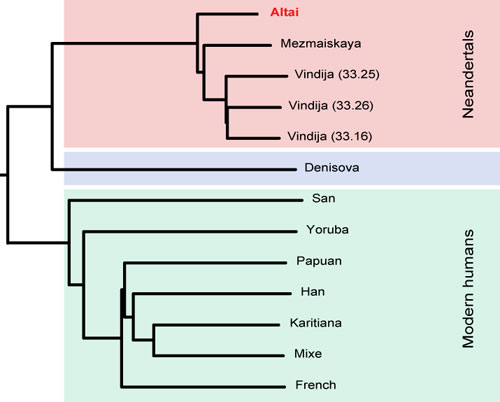

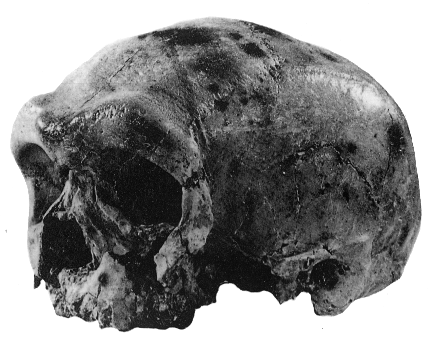









 and a finger bone show the Denisovans were a distinct group.")







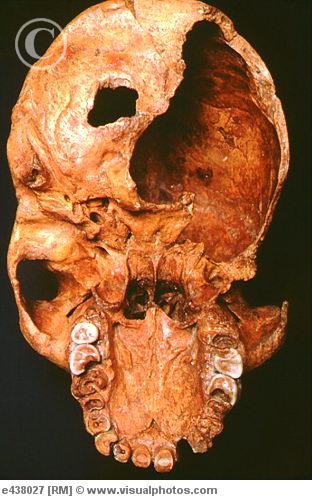





















.")































































































 Jonathan Bloch, één van de onderzoekers, laat hier het schedeltje en afgietsel van de herseninhoud van de primitieve aap zien. Op het scherm op de achtergrond is het virtuele 3D-model zichtbaar.
Jonathan Bloch, één van de onderzoekers, laat hier het schedeltje en afgietsel van de herseninhoud van de primitieve aap zien. Op het scherm op de achtergrond is het virtuele 3D-model zichtbaar. 


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
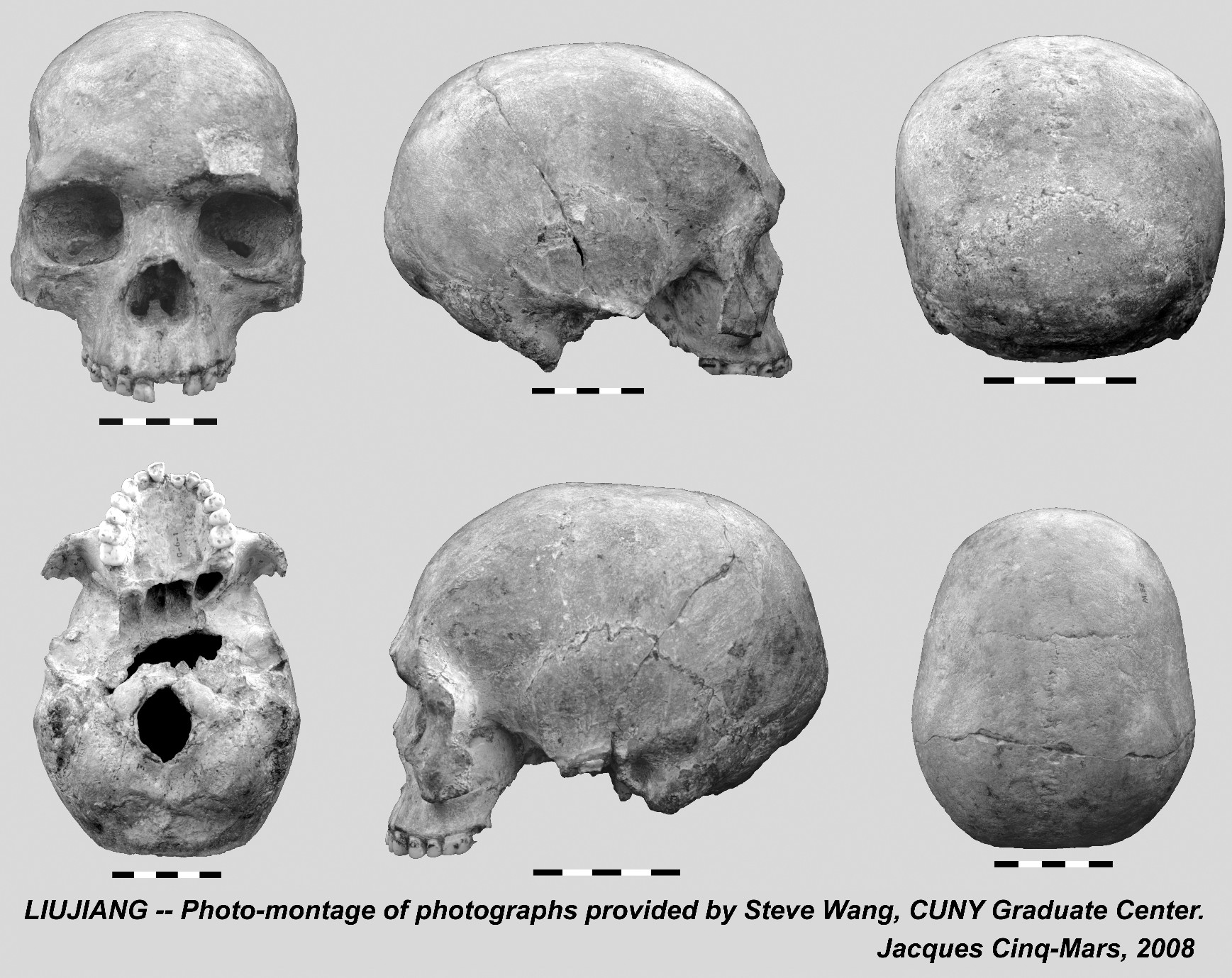{kind=link}
{kind=link}
{kind=link}
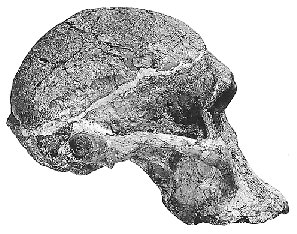{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
( March 6, 2009
March 6, 2009
” a trackmaker assignment is a hypothesis.
Eubrontes isn’t a dinosaur body-fossil genus. It’s an ichnotaxon–in this case a footprint genus. There are no Eubrontes skeletons out there. The name is restricted to the footprint morphotype alone. Ignoring the issues with slapping Latin bionomials on sedimentary structures, this is a common practice and Eubrontes fossils represent a fairly characteristic type of footprint shape found across North America in Lower Jurassic rocks.
The question of what animal made Eubrontes is a different issue, but identifying a footprint as a Eubrontes track is a matter of studying the footprint itself and comparing it to other footprints. There need not be dinosaur skeletal feet preserved nearby that fit the tracks. Again, the issue of who made the fooprint is a different issue from figuring out whether or not the track morphology is consistent with the morphotype that is known as Eubrontes. ”
http://scienceblogs.com/pharyngula/2009/03/06/how-did-dinosaurs-sit-down/