KIEUWBOGEN en creationisten
oktober 17, 2012 1 reactie
INHOUD GLOS E →
Kieuwbogen en creationisme
zie –> EMBRYOLOGY development biology
door renefransen jun.08, 2012,
De kieuwboog (of kieuwspleet) is een kenmerk van het ontwikkelende embryo van alle gewervelden. Een serie plooien in de ‘nek’ die bij verschillende dieren zich op verschillende wijze ontwikkelen. Bij de mens leveren ze onder meer de onderkaak, trommelvlies en gehoorbeetjes op.
Over de kieuwspleten is nogal wat te doen. Is het een bewijs voor evolutie? Een bewijs dat mensen van vissen afstammen? Recapituleren wij onze vissige afkomst ermee in de embryonale ontwikkeling?
Creationisten hebben nogal wat kritiek op de suggestie dat het voorkomen van kieuwbogen bij alle gewervelden wijst op gemeenschappelijke afstamming. Een deel van die kritiek, met een inhoudelijke weerlegging ervan, staat op de website ‘Playing chess with pigeons’ van Troy Britain.
Het stuk is behoorlijk goed leesbaar en geeft een mooi overzicht van wat kieuwbogen wel en niet zijn en vooral: hoe er in het verleden (voor Darwin/ en ook door Darwin ) over geschreven is.
Britain pakt een aantal door creationisten geponeerde stellingen aan en weerlegt hun argumenten overtuigend.
Er blijven ongetwijfeld argumenten onbesproken, maar het artikel is een aanrader.
Het artikel is voorzien van een grote hoeveelheid links, dus alles is na te checken.
(Hieronder mijn aangevulde (= tsjok) voorlopige vertaling van het artikel van Troy Britain )

“Gill slits” by any other name…
Andere namen en dergelijke voor ”Kieuwbogen ” ….
Charles Darwin zei ooit dat ; het bewijs uit de vergelijkende anatomie van embryo’s ”veruit de sterkste klasse van feiten” is ,ter onderbouwing van de theorie van de gemeenschappelijke afstamming (Darwin, 1860) en dacht___ alhoewel het sindsdien is overschaduwd door de genetica___dat het een van de meest dwingende deelverzamelingen van bewijs voor evolutie, zal blijven .
En misschien zijn wel de meest opvallende details van de vergelijkende embryologie bij gewervelde dieren, de structuren die in de volksmond bekend staan als“kieuwspleten”.
Embryonale “kieuwspleten” of “branchiale kloven” (branchia is Grieks voor kieuw) of (beter )faryngale spleten (groeven, plooien, enz.) maken deel uit van wat genoemd wordt het ”faryngale apparaat ” dat zich ventraal en lateraal onder het hoofd / in nek en halsgebied , van alle gewervelde dieren bevindt en wél tijdens de “ pharyngula fase “van de(embryonale) ontwikkeling.
Zowel in “vissen”, en de larve van de amfibieën, ontwikkelen deze groeven zich tot uitstekende (kieuw-matige )ademhalingsorganen gebruikt om zuurstof uit water te halen, terwijl in amnioten (“reptielen”, vogels en zoogdieren) deze structuren zich ontwikkelen in andere inwendige organen / structuren en functies .
Voordat ik verder ga, volgt hier eerst een korte uitweiding over de “vissen”.
In dit artikel zal ik regelmatig gebruik maken van het woordje ”vis” in de algemene betekenis, maar er dient meteen te worden opgemerkt dat de term zoals die vaak wordt gebruikt ___ , en daarbij verwijst naar alle gewervelden die zwemmen in het water, en vinnen en kieuwen bezit _____ geen geldige wetenschappelijke classificatie is .
Dit komt omdat de drie belangrijkste soorten van dieren die in de volksmond “vis” noemen ; -de Chondrichthyes (haaien, roggen en chimaeras ), de Actinopterygii (de meerderheid van levende vissen ), en de Sarcopterygii (kwastvinnigen , de groep waaruit viervoetige landdieren, dat wil zeggen de tetrapoden , evolueerde)-___ geen monofyletische groep vormen .
Ze zijn ook niet erg nauw verwant aan elkaar, ( = ze volgen al zeer lang gescheiden evolutiepaden ) ondanks een aantal gemeenschappelijke uiterlijke overeenkomsten (zoals kieuwen).
Zo delen bijvoorbeeld de extante Sarcopterygii-, long-vis en coelacanthen een meer recente gemeenschappelijke voorouder met ons (en alle tetrapoden) dan met de andere ”vissen”.
OK, Laten we na deze preciseringen verder gaan met ons eigenlijke verhaal ….
De “keelholte uitrusting ” bestaat uit een reeks gekoppelde pharynxbogen en scheuren die op de buitenzijde ontwikkelen met een overeenkomstige reekskeelholte zakjes aan de binnenzijde van de keel, gescheiden van de externe spleten door een vlies (meer details hieronder )).
In feite is het bezit van deze structuren op een bepaald punt in de embryonale ontwikkeling, samen met een holle dorsale zenuw koord, een notochorda en eenpost anale staart, een definiërend kenmerk van het phylum Chordata waar wij en alle andere gewervelde dieren toe behoren.

Bovenstaande illustratie is schematisch en niet bedoeld om fotografisch nauwkeurig te zijn ( ik wens namelijk niet beschuldigd te worden door creationisten van het “transporteren” en overnemen van een “frauduleuze “illustratie). Hieronder vindt u daadwerkele actuele foto’s van zowel een roggen –(skate) embryo als van een menselijk embryo ter vergelijking.
De kieuw- structuren in de embryo’s van elasmobranchia (vissen-onderverdeling van Chondrichthyes die haaien, roggen en chimerae(draakvissen) omvat )is ook veel minder afgeleid/veranderd dat in andere verder ontwikkelde (been) “vissen” en dus over het algemeen beter vergelijkbaar met de embryo’s van amnioten dan de overeenkomstige structuren in de benige “vissen” (die aanzienlijk zijn gewijzigd).
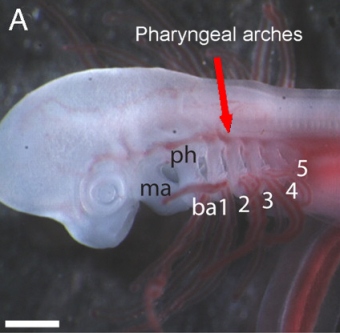

De eerste van de bogen, de onderkaak boog, vormt de kaak in Gnathostome gewervelde dieren (Gnathostomes ).
De meeste gewervelde dieren ontwikkelen een totaal van zes bogen, maar de volledige set wordt meestal alleen behouden tot in de volwassenheid door hexanchiforme haaien.
Hexanchiformes zijn zeer plesiomorf ; wat betekent dat ze meer lijken op vroegere soorten haaien. Sommige soorten hexanchiformes ontwikkelen nog een zevende boog. Ook de extante primitieve kaakloze gewervelde, de lamprei , bezit nog zeven kieuwopeningen.
In menselijke embryo’s zijn de vier bogen zichtbaar en worden gevormd rond de 4e week van de ontwikkeling , de 5e boog is van voorbijgaande aard, terwijl de 4e en 6e versmelten tot één. En zoals bij de meeste gewervelde landdieren ,zijn ze gewijzigd tijdens de embryonale ontwikkeling in niet-kieuw -structuren in de volwassene.
De standaard evolutionaire verklaring van de vorming van de faryngeale apparaat in de embryo’s van amnioten is natuurlijk dat hun verre voorouders waren in feite in het water levende, Gill lager, organismen en dat de restanten van de embryonale kieuw-achtige structuren zijn bewaard gebleven in hun embryonale ontwikkeling. En hoewel deze structuren in de embryo’s van amnioten zich nooit ontwikkelen tot de werkelijke kieuwen, zijn ze in het verleden is informeel aangeduid als “kieuwspleten” als gevolg van hun algemene anatomische overeenkomst met de bijbehorende structuren van het water levende gewervelde dieren die wel ontwikkelen tot kieuwen.
Zoals je misschien voorstellen dat het feit dat de zogenaamde “kieuwspleten” zijn te vinden in amnioten embryo’s is een grote bugaboo voor creationisten, rijden ze een hoop energie te steken door de jaren heen in het minimaliseren van de gelijkenis van deze structuren (Price, 1924), en in kaak iedere wetenschapper die praat over hen.
Dit heeft geleid tot de ontwikkeling van een vrij standaard standaardtekst over het onderwerp onder de creationisten. Hier zijn enkele goede voorbeelden, de eerste wezen van Drs. Elisabeth & Tommy Mitchell , die van de jonge aarde creationist organisatie Answers in Genesis en zijn beide schijnbaar artsen (een is zelfs verondersteld een verloskundige zijn):
De zogenaamde kieuwspleten van een menselijk embryo hebben niets te maken met kieuwen, en het menselijke embryo wordt niet door een vis podium of een ander evolutionair stadium. De ontwikkeling van het menselijk embryo onthult gestage vooruitgang in de richting van een volledig functionele menselijk lichaam. Nooit in de loop van de ontwikkeling van een menselijk embryo wordt zuurstof uit het water te absorberen als vis doen met kieuwen. (Het menselijk embryo is volledig voorzien van zuurstof via de navelstreng.) In feite zijn deze ‘kieuwspleten’ zijn niet eens spleten.
Dus wat zijn deze ten onrechte zo genoemde structuren? Eigenlijk zijn ze niets meer dan plooien in de regio van de keel van de kleine embryo’s. Door de 28ste dag van het leven, het embryo de hersenen en het ruggenmerg lijken van te voren te racen van de rest van het lichaam van de groei. Daarom is voor een tijd, het ruggenmerg is eigenlijk langer dan het lichaam, waardoor het lichaam te krullen en het buigen van de nek naar voren. (Dit gekruld embryo met de lange ruggenmerg wordt ten onrechte beschuldigd door sommige mensen van het hebben van een staart.) Net als veel mensen een dubbele kin te ontwikkelen bij het buigen van de nek naar voren, zodat het embryo plooien in haar nek als gevolg van deze buiging heeft.
. […] De buiten-en middenoor en de botten, spieren, zenuwen en klieren van de nek ontwikkelen zich uit deze vouwen slechts oppervlakkig ooit deze belangrijke plooien niet lijken op kieuwen, de faryngeale bogen zijn niet meer te maken met kieuwen dan sterren zijn tot straatverlichting. (Mitchell & Mitchell 2007, mijn cursivering)
Hier hebben we “creationistische anatoom” en veteraan antievolution krijger, Dr David Menton , ook vanAnswers in Genesis:
Menselijke embryo’s “kieuwspleten” Op een bepaald moment van hun ontwikkeling BEWIJZEN dat de mens evolueerde VAN WATER-, VIS EN amfibie, VOOROUDERS.
Je kan nauwelijks de middelbare school of hogeschool nu dagen bij te wonen met uit het horen of lezen van deze “kanjer”. In feite zijn noch kieuwen noch hun spleten vinden op elk stadium van de embryonale ontwikkeling van een zoogdier waaronder de mens. De plooien in de nek van de zoogdieren embryo, die ten onrechte worden “kieuwen”, zijn niet kieuwen in de betekenis van het woord en nooit iets te maken met de ademhaling. Ze zijn slechts flexie vouwen of rimpels, in de nek regio als gevolg van de sterk gedraaid hoofd en uitstekende hart van de zich ontwikkelende embryo. Deze plooien uiteindelijk ontwikkelen tot een deel van het gezicht, binnenoor, de amandelen, de bijschildklier en thymus. Geen gerenommeerde medische embryologie tekst stelt dat er ‘kieuwspleten’ in zoogdieren.
Toch is de kieuw spleet mythe bestendigd in vele middelbare school en universiteit biologie tekstboeken als “wetenschappelijk bewijs” voor evolutie. Zelfs Dr Spock in ‘Baby and Child Care’ zijn boek beweert dat “als de baby ligt in het vruchtwater van de baarmoeder, hij kieuwen zoals een vis heeft.” Misschien is de “gill slit” mythe nog steeds worden onderwezen, omdat er geen beter “bewijs” voor evolutie. Hoeveel van jullie zijn de kieuw gleuf mythe geleerd op school??(Menton 1991, mijn cursivering)
Vervolgens hebben we ‘intelligent design’ creationist dr. Cornelius Hunter , een biofysici en Fellow aan hetDiscovery Institute Center voor de vernieuwing van Wetenschap en Cultuur :
Nu, wat is er mis met Coyne’s [bioloog Jerry Coyne (2009)] beschrijving van de ontwikkeling van vertebraten? Ten eerste gewervelde dieren gewoon niet beginnen met de ontwikkeling op zoek als embryonale vis. Dit is wat de evolutietheorie voorspelt, en wat evolutionisten willen zien. Ja er zijn overeenkomsten, maar dit is nog een ander geval van theorie-gedreven, in plaats van data-driven, denken.
[…] Vervolgens Coyne begint meteen te verwijzen naar de groeven tussen de kieuwbogen, in menselijke embryo’s, zoals kieuwspleten. Maar mensen hebben geen kieuwen als volwassenen. De mens nooit kieuwen op elk moment te hebben. Er is dus geen basis om te verwijzen naar de groeven als “kieuwspleten” afgezien van de domme evolutionaire mandaat dat de kieuwbogen zijn een evolutionair overblijfsel dat vandaag de dag net gebeuren om structuren te vormen, zoals het middenoor, het strottenhoofd, buis van Eustachius, en bloedvaten en zenuwen.
Dus Coyne interpreteert het bewijs volgens de theorie die hij denkt dat waar is, en dan presenteert de belachelijke interpretatie zo krachtig bewijs voor de theorie. Ik zou verbaasd zijn geweest als ik niet had gezien zoals een cirkelredenering zo vaak eerder in de evolutie genre.(Hunter 2010, e MphasiS mijne)
Tot slot hebben we nog een intelligent ontwerp creationist, Dr Jonathan Wells een Senior Fellow van het Discovery Center Instituut voor de vernieuwing van Wetenschap en Cultuur, en ogenschijnlijk een ontwikkelingsstoornis bioloog:
Halverwege ontwikkeling, alle gewervelde embryo’s bezitten een reeks van plooien in de nek, of keelholte. De convexe delen zijn genoemd faryngeale “bogen” of “kammen,” en de holle delen worden genoemd faryngeale “kloven” of “zakken.” Maar faryngeale plooien zijn niet kieuwen. Ze zijn niet eens kieuwen in pharyngula-stage vis embryo’s.
In een vis, faryngeale plooien later ontwikkelen tot kieuwen, maar in een reptiel, zoogdier, vogel of ze zich ontwikkelen tot andere structuren geheel (zoals het binnenoor en bijschildklier). In reptielen, zoogdieren en vogels, keelholte plooien zijn zelfs nog nooit rudimentaire kieuwen, ze zijn nooit ”gill-achtige”, behalve in de oppervlakkige zin dat ze een aantal evenwijdige lijnen in de nek regio te vormen.
[…] Met andere woorden, er is geen reden om embryonale faryngeale zakjes noemen “kieuw-achtige.” De enige rechtvaardiging voor die term is de theoretische bewering dat zoogdieren zich ontwikkeld van vis-achtige voorouders.
[…] De enige manier om “kieuw-achtige” structuren te zien in menselijke embryo’s is aan de evolutie lezen in ontwikkeling. (Wells, 2000, pp.105-106, mijn cursivering)
Er zijn dus blijkbaar op zijn minst drie belangrijke punten van de creationisten willen maken:
- Faryngeale structuren van amniote embryo’s nooit functioneren als kieuwen en moet daarom niet worden aangeduid als “kieuwspleten”.
- Wat lijkt op de kieuwen van de aquatische gewervelde dieren de faryngeale structuren van amnioten heeft, is het oppervlakkig.
- Het zien van de faryngeale structuren van amniote embryo’s als kieuw-achtige en noemde ze kieuw-spleten ondanks hun niet functioneren als kieuwen is “het lezen van de evolutie in de ontwikkeling”.
Uiteraard is er enige overlap is er maar laten we ons elke te onderzoeken op hun beurt.
Nummer 1: Gill spleten door een andere naam …
Dit is in wezen een semantische discussie over wat we noemen deze structuren in gewervelde embryo’s in combinatie met een stroman argument en deze combinatie is verre van nieuw.
Als je alleen creationistische bronnen te lezen over dit onderwerp kunt weg zou komen met de sterke indruk dat de controversiële midden van de 19e eeuwse Duitse bioloog Ernst Haeckel (1834-1919), het idee dat amniote embryo’s “kieuwspleten” out of geheel hebben verzonnen doek, ergens in de jaren 1860, om overeind te houden Darwin’s niet en ongegrond idee van evolutie (Morris 1989) (Grigg 1996) (Harrub 2001) (Butt 2010) (Sherwin 2011).
Maar de waarheid is dat deze structuren werden oorspronkelijk beschreven bij vogels en zoogdieren embryo’s in 1825 door de Duitse anatoom / embryoloog Martin Heinrich Rathke (1793-1860) (Rathke 1825a & 1825b). Rathke genoemd hen beiden als “Schlundspalten” (“keel kloven” ) en “Kiemenspalten” (“gill kloven”), wat betekent dat, door de moderne creationistische schatting, zijn ze verkeerd benoemd zijn vanaf moment van hun ontdekking (Tuttle 1884, pp 111-112 ) (zie ook Ascherson 1848). En de moderne creationisten waren niet de eerste om dergelijke semantische bezwaar te maken:
De ontdekking en ware interpretatie van de branchial scheuren in het embryo van de hogere gewervelde dieren behoort om te Rathke, […] Kort na deze, Huschke illustreerde de zaak, met name in het kuiken […]. Zeer recent Reichert heeft gevoerd het onderwerp diep, hij noemt de kieuwbogen viscerale bogen-Müller Archiv für 1837 zijn bewering, dat deze niet zijn kieuwbogen,is slechts een geschil over een woord. Werd nooit gedacht dat de onderdelen in kwestie juistekieuwen, maar ze zijn vasculaire bogen, die in alle opzichten analoog aan de vasculaire bogen van de kieuwen van vissen, niet alleen vertakking als deze (Wagner 1844, blz. 111,mijn cursivering.).
Wat Rudolf Wagner zei 166 jaar geleden (let op de datum) geldt net zo waar vandaag de dag, niemand beweert dat ‘kieuwspleten’ ooit functie als kieuwen in amnioten. Dat creationisten steeds harp op dit eenvoudige stroman bashing niet als een wetenschapper, dat ik ben me bewust van, behalve een-en bespreek ik hem later-heeft sinds Wagner schreef die verklaring, beweerde dat zij doen.
Dus dat laat ons achter met de semantische bezwaar tegen het informele gebruik van de term kieuwspleten te verwijzen naar deze structuren, dat is een beetje als het krijgen werkte up over het aanroepen van de mens stuit een ‘stuitje’, omdat het nooit werkt als een echte staart bij de mens .
Oh wacht, ze waarschijnlijk niet graag dat ook niet.
Een beetje meer serieus, wat dacht je van de kleine rudimentaire vleugel knoppen van kiwi’s (geslachtApteryx; kleine vliegende vogels gevonden in Nieuw-Zeeland)? Ze zijn duidelijk niet gewend om te vliegen op elk punt in het leven van een kiwi. Is het ongepast te worden genoemd als vleugels?
Of wat dacht je van iets een beetje dichter bij het onderwerp bij de hand, de achterste ledematen in de embryo’s van walvisachtigen (walvissen en dolfijnen)? In walvisachtigen ze meestal niet ontwikkelen tot achterpoten en worden gewoon weer opgenomen in het lichaam. Af en toe walvissen en dolfijnen worden geboren met externe achterpoot structuren maar deze zijn niet meestal gekoppeld, symmetrisch, en goed gevormd. Maar ze zich voordoen en ontwikkelen ze recht werden ze zou moeten zijn als walvissen zouden achterpoten groeien.

Copyright 2006, dr. JGM Thewissen
Deze structuren zullen nooit worden gebruikt om nog lopen of zwemmen, dus moet vergelijkende ontwikkelingsbiologie teksten worden verboden om de term ‘achterpoot bud “te gebruiken bij de verwijzing naar deze structuren in walvisachtigen?
Natuurlijk niet.
We kunnen, informeel, bel faryngeale spleten “kieuwspleten” op dezelfde manier noemen we kiwi ‘vleugels’, vleugels of walvisachtigen “ledematen”, ledematen, want ondanks het feit dat ze niet meer ontwikkelen tot functionerende vleugels, ledematen of lamellen in de organismen die ze draagt, omdat zij duidelijkhomoloog die tekens organismen ze blijven deze functies.
De creationistische bezwaar dat het verwijzen naar faryngeale structuren als kieuwspleten veronderstelt evolutie (punt # 3), indien gegrond, ook zou gelden voor kiwi vleugels en de walvis achterste ledematen sinds de grote onderscheid tussen deze verschillende homologe structuren, vanuit een evolutionair oogpunt, slechts tijd. Echter zal men tevergeefs zoeken naar websites boos kaak stellen van het gebruik van de term “vleugels” of “ledematen” om die structuren te beschrijven te vinden. [Natuurlijk nu een aantal joker zal een uurtje Google-mining alleen maar om de een tin-foil hat drager die er zijn dat heeft dit bezwaar alleen maar zodat ze kunnen gooi het in mijn gezicht te vinden.]
Ik stel voor dat de reden waarom we niet zien creationisten het maken van een stank over deze andere voorwaarden is dat ze niet raken bij mensen de manier waarop de aanwezigheid van “kieuwspleten” in amniote embryo’s heeft (gezien het feit dat we amnioten). En het behoud van de mens als speciale goddelijke creaties hun primaire (theologische) zorg.
“Wacht even!”, Verzetten de creationisten, “Je bent nummer drie doet de hele tijd!”.
Heb geduld, ik doe dat.
Nummer 2: Zijn “kieuwspleten”, spleten? En zijn de overeenkomsten tussen kieuwspleten en faryngeale structuren oppervlakkig?
Ongeacht van wat we noemen deze faryngeale structuren in amniote embryo’s, ze dragen in feite alle overeenkomsten met de kieuwen van de “vis”, of in ieder geval de faryngeale structuren in “vis” embryo’sdie worden kieuwen? Verklaringen van creationisten zoals die hierboven aangehaalde geven de indruk dat faryngeale bogen en kloven waren weinig meer dan “flexie plooien” of “evenwijdige lijnen” in de ectoderm(wat wordt de huid onder andere) van het embryo, zoals de ruggen je zou kunnen krijgen in een tuinslang die is geknikt.
Maar vraag jezelf dit, als dat alles is ze ook waren, als ze echt net als een geknikte slang of een dubbele kin, is het waarschijnlijk dat de biologen zo zou zijn onder de indruk van deze structuren dat zij zouden overwegen hen een van de bepalende kenmerken van het phylum Chordata? Zoals gebruikelijk, het verhaal is veel complexer dan creationisten laten merken.
Het feit is, als we kijken naar de details van deze structuren, vinden we niet slechts een oppervlakkige gelijkenis, maar eerder een verzameling van anatomische overeenkomsten tussen de kieuwbogen van de “vis” en de embryonale faryngeale structuren van amnioten.
De faryngeale bogen, waarin elk van de gespleten / zakken voorafgaan, zijn een stuk meer dan eenvoudige maal richels in de huid omdat ze bevatten een hele reeks elementen, skelet, spieren, neurale en de bloedsomloop. De kern van elk van de bogen een kraakbeenhoudend element geflankeerd door een aorta (bloedvat) schedelzenuwen en spier elementen.
Hier is een illustratie dat ik greep uit de ‘interwebs’ en aangepast aan de relaties van de verschillende elementen in de faryngeale structuren van gewervelde embryo’s laten zien:

Bron bionalogy.com, met wijzigingen.
Spleten & Pouches
Creationisten zijn dol op te herhalen dat in amnioten, “kieuwspleten” zijn niet alleen niet kieuwen, ze zijn niet eens spleten. En ze gedeeltelijk corrigeren indien de “spleet” een een onbelemmerd opening van de buitenkant van de nek van de in de keel van zoogdier embryo’s. Dat wil zeggen zij technisch corrigeren zoveel normale zoogdier ontwikkeling gaat, maar dit is niet het geval alle niet-”vis” vertebraten, ook niet altijd het geval zoogdieren met inbegrip van mensen.
In de meeste ‘normale’ amnioten, inclusief wij mensen, het enige wat houdt ons van het hebben van ten minste een open ‘sleuf’ in onze eerste faryngeale spleten zijn de dunne membranen van de huid, hettrommelvlies is, of trommelvliezen. Zonder uw trommelvliezen zou je open kanalen van je uiterlijke oren, door je midden oren en buis van Eustachius , in je keel.

Net als voor andere faryngeale spleten, een aantal van deze nog volledig open in de larve van sommigeamfibieën die ademen via de kieuwen net als “vis” te doen. Verder is de “post oor” faryngeale spleten ook tijdelijk te openen in een aantal “reptielen” en vogels opnieuw te sluiten in de loop van de normale ontwikkeling.
Dus in het geval van sommige amfibie zijn kieuwspleten en enkele “reptielen” en vogels zij feitelijk sleuven (openingen in de keel).
En hoewel het waar is dat zoogdieren (inclusief mensen) normaal gesproken niet letterlijk ‘spleten’ (openingen) bezitten in de nek, af en toe mensen met een abnormale ontwikkeling te doen. Er zijn gevallen geweest waar mensen zijn geboren, waar, bijvoorbeeld, een of beide van hun tweede faryngeale spleten perforeert en blijft open na hun geboorte (zie de voorbeelden hier , hier en hier ). Dit fenomeen ( atavisme), is vergelijkbaar met de gevallen waarin walvissen geboren met achterpoten.
Pharynxbogen
Elke keelholte boog sport een kraakbeen bar, de eerste (mandibulaire) boog, zoals eerder werd opgemerkt, ontwikkelt zich tot de kaak beenderen van alle jawed gewervelde dieren. In “reptielen” en vogels is er sprake van meerdere botten van de onderkaak, terwijl bij zoogdieren het zich ontwikkelt in het enkelvoud onderkaak bot (de onderkaak ) en twee van de botten van het middenoor (de incus en de Malleus ). Zie: “Evolutie van zoogdieren gehoorbeentjes ”
Figure 1.4.3. A comparison of the jawbones and ear-bones of several transitional forms in the evolution of mammals. Approximate stratigraphic ranges of the various taxa are indicated at the far left (more recent on top). The left column of jawbones shows the view of the left jawbone from the inside of the mouth. The right column is the view of the right jawbone from the right side (outside of the skull). As in Figure 1.4.1, the quadrate (mammalian anvil or incus) is in turquoise, the articular (mammalian hammer or malleus) is in yellow, and the angular (mammalian tympanic annulus) is in pink. For clarity, the teeth are not shown, and the squamosal upper jawbone is omitted (it replaces the quadrate in the mammalian jaw joint, and forms part of the jaw joint in advanced cynodonts and Morganucodon). Q = quadrate, Ar = articular, An = angular, I = incus (anvil), Ma = malleus (hammer), Ty = tympanic annulus, D = dentary. (Reproduced from Kardong 2002, pp. 274, with permission from the publisher, Copyright © 2002 McGraw-Hill)
Voor meer informatie over de evolutie van de zoogdieren kaak en oor botten, zie:
“Tussenvormen en overgangsvormen – “(1999-2012) door Douglas Theobald http://www.talkorigins.org/faqs/comdesc/section1.html
http://www.talkorigins.org/faqs/comdesc/section2.html
- “ Een blijven achtervolgen van Jaw”(1990) door Stephen Jay Gould
- “ Evolutie van zoogdieren gehoorbeentjes ”(Wikipedia)
In “vis” de andere bogen ondersteunen de kieuwen, terwijl in amniote ze worden gewijzigd in andere structuren in de nek. Bijvoorbeeld de tweede boog zich ontwikkelt tot de tongbeen in de keel en destijgbeugel in het oor van zoogdieren.

Arches kleur legende: 1e = I, 2e = II, 3e = III, IV = 4e, 6e = VI
Aorta-bogen
Evenwijdig aan elkaar lopende kraakbeenachtige bar in de pharynxbogen worden slagaders genoemd aorta bogen . In “vis” en amfibieën larven ze route bloed uit het hart via de kieuwen (waar het dumpt CO 2 en hoger O 2 halen) en naar de dorsale aorta.
Ook de aorta bogen van amniote embryo’s route bloed uit het hart rond de nek tot aan de dorsale aorta.Maar in tegenstelling tot de “vis” en amfibieën zij nooit de ontwikkeling van de fijne haarvaten en Gill filamenten van de juiste kieuwen. In plaats daarvan worden gewijzigd tijdens de ontwikkeling voor andere doeleinden. Bijvoorbeeld in tetrapoden (amfibieën, “reptielen”, vogels en zoogdieren) de 6e aortaboog ontwikkelt zich tot de longslagader die verbinding maakt met de longen. Interessant is dat deze zelfde regeling is toevallig te vinden in de aorta bogen van bestaande longvissen ( dipnoans ) (Kardong 2009, p.456), die op basis van vergelijkende anatomie en genetica (Venkatesh et al. 2001) (Takezaki et al.. 2004) maken deel uit van de groep van de “vis”, Sarcopterygii, die, zoals ik in het begin, is de groep van “vis” meest verwant aan de tetrapoden.

Van Eastern Kentucky University BIO 342 Vergelijkende gewervelde Anatomie site, met wijzigingen (ingekleurd en bestempeld als de harten).
Let op de relatieve positie van het hart van de pharynxbogen in het schema van amniote embryo bovenstaande, net onder en aan de achterzijde van de aorta bogen. In “vis” de ontwikkeling van het hart onderhoudt deze relatieve positie van de faryngeale structuren-vlak onder ( ventraal ) en naar achteren (posterior ) van de kieuwen. In amnioten echter de faryngeale structuren van amniote embryo’s uitgroeien tot de kaak en de nek en het hart raakt gescheiden van de nek van het lichaam in de volwassen (dit is van belang als we kijken naar de volgende zenuwen), naar beneden in de borst.
(Stansfield 1977)
Zenuwen
De eerste faryngeale boog, die zoals reeds vermeld, ontwikkelt zich tot de kaak beenderen in allegnathostomes (kaken gewervelde dieren), die wordt geïnnerveerd door de 5 e hersenzenuw (de trigeminus), die de hersenen verbindt met de kaken en delen van het gezicht.

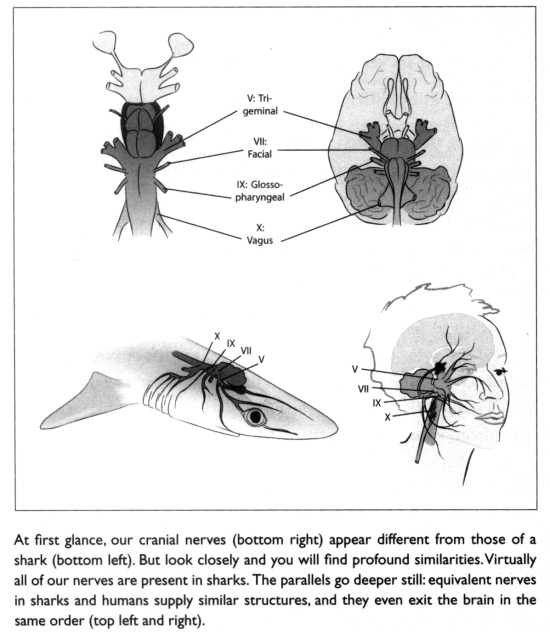
(Shubin 2009, p.92)
Een andere zenuw van belang is 10e craniale of nervus vagus . In de “vis” dit komt uit de basis van de hersenen en maakt verbinding met verschillende organen in het lichaam zo nu en dan sturen ze takken af langs de weg. Een van deze takken, de 4e, innerveert de kieuwen van de 6e faryngeale boog in “vis”, en dat is een mooie rechte pad vanaf de bovenkant (dorsale) deel van de “vis” aan de onderkant (ventrale) deel (zie het plaatje hieronder).
 Coyne 2009, p.83
Coyne 2009, p.83
Echter, zoals we eerder zagen het bij zoogdieren de 4e en 6e pharynxbogen zekering-na de tijdelijke 5e boog wordt weer opgenomen. De 4th/6th boog gewijzigd door ontwikkeling draagt dan de vorming van het strottenhoofd. De truc is dat wanneer de verlengde nek geëvolueerd amnioten en de 4e tak van de nervus vagus-nu de laryngeus recurrens zenuw -raakte verslaafd door de 6e aortaboog en naar beneden getrokken in de borst, het veranderen van wat was een vrij rechte schot langs de 6e kieuwboog in “vis”, in een omweg, waarbij de nervus vagus komen uit de basis van de hersenen reist naar beneden in de borst, lus rond de longslagader en gaat dan weer de nek in het strottenhoofd.
Dit heeft geleid tot een laryngeale zenuw van komische proporties in de giraffen.

Als je niet preuts en u wilt dit zelf zien , de uitstekende Britse tv-show Inside Nature’s Giants deed een dissectie van een giraffe in de 4e aflevering , met de nadruk haar laryngeus recurrens zenuw, ik kan het niet aanraden dit programma sterk genoeg.
Misschien is het niet verwonderlijk is het dan, dat giraffen geen stem geven heel veel. Tegen de tijd dat het signaal om een geluid te maken reist van hun hersenen in hun nek en weer omhoog ze vergeten wat ze geluiden maakt over!
In ieder geval is er geen voor de hand liggende zin om deze stand van zaken van een opnieuw ontworpen, engineering oogpunt, maar het is volkomen logisch als amnioten zijn opportunistisch gewijzigd “vis”.
Fysiologie, genetica
Vergelijkingen tussen faryngeale bogen in de “vis” en amnioten verder reiken dan hun bruto anatomie, er zijn ook gedeeld fysiologische en genetische overeenkomsten.
For example, while it is universally admitted that amniote embryos don’t use their “gill slits” to breath, there is at least one other physiological function originating in the pharyngeal apparatus that is conserved between “fish” and amniotes; regulating calcium salts (a major component of vertebrate bones). “Fish” regulate the amount of calcium in their bodies using their gills. Amniotes do this by secreting hormones from their parathyroid glands which just happens to develop from the 3 rd and/or 4 th embryonic pharyngeal pouches (Okabe & Graham 2004).
For more see:
At the genetic level there are two significant families of genes (Hox & Dlx) which control the development of the pharyngeal region of all vertebrates.

Kuratani 2004, p. 337 (fig. 3)
A nested set of Hox genes control the development of the pharyngeal structures head to tail (anterior to posterior) from just after the first pharyngeal arch (Hunt et al. 1991) (Prince et al. 1998) (Kuratani 2004). And likewise a nested set of Dlx gene are expressed in the development of the pharyngeal structures in the front to back (ventral/dorsal) direction (Schilling 2003) (MacDonald et al., 2010).
Paleontology bonus section
It addition to all the interesting stuff above, the pattern of vertebrate embryological development—wherein an aquatic body plan is modified into one better adapted to a terrestrial environment—roughly matches that of the fossil record (“fish” first, amphibians and tetrapods later).
De eerste chordadieren te verschijnen in het fossielenbestand waren de cephalochordates , kleine vis-achtige organismen die beide kaken en waar het hoofd ontbrak, net als de levende Branchiostoma die zich voedt door het filteren van water door de kieuwen. Daarna chordadieren verschijnen in het fossielenbestand in een bepaald patroon (let op dat deze zeer wordt vereenvoudigd):
- Chordates with true heads ( Craniates) but remained jaw-less similar to the living hagfish.
- Basal jawless vertebrates ( Agnathans) similar to the living lamprey.
- The larvae of lamprey ( ammocoetes ) physically resemble Branchiostoma and filter feed via their gills in a similar manner.
- Various “fishes” with jaws ( Gnathostomes ) .
- The jaws of some early Gnathostomes ( Acanthodians ) resemble enlarged gill arches.
- Tetrapod like fish ( Tetrapodomorpha , a sub group of the Sarcopterygiimentioned earlier).
- Basal tetrapods(amphibian-like animals).
- Some early tetrapods retained internal gills like Tetrapodomorpha fish.
- Basale amnioten ( Anapsid”reptielen”)
- Basale Synapsids (de voorouders van de zoogdieren) en Diapsids(de voorouders van de “reptielen” en vogels).
- Basale zoogdieren en vogels.
- Basale zoogdieren gelegd eieren veel op die van diapsids; monotreme zoogdieren nog steeds doen.
[Let op: het tijdspatroon van de verandering besteld in het fossielenbestand, dat wil zeggen fauna successie , was een vaststaand feit vóór Darwin gepubliceerd On the Origin of Species (1859). Een feit die door geologen die zich in de eerste plaats creationist in de vooruitzichten.]
Op dit punt, ik hoop, ik heb aangetoond dat de creationistische karakterisering van de faryngeale structuren van amniote embryo’s als zijnde slechts oppervlakkig kieuw-achtige “vouwen” of “grooves”, is voorbij misleidend. Er zijn tal van anatomische overeenkomsten, ten minste een gedeelde fysiologisch proces (dat is niet de ademhaling), en een gedetailleerde onderliggende genetische gelijkenis als goed, die geen van alle creationisten zelfs proberen om een coherente uitleg te geven voor.
Ten minste drie van de vijf creationisten (de Elizabeth Mitchell, Menton en Wells) hierboven geciteerd had moeten zijn, op basis van hun opleiding (een verloskundige, een anatoom en een ontwikkelingsstoornis bioloog), op de hoogte van de anatomische / embryologische feiten die ik hierboven gedocumenteerd. Toch is en zich niet een van deze informatie aan hun lezers ze in plaats daarvan gekozen om hen te vertellen dat faryngeale spleten in amniote embryo’s zijn slechts oppervlakkige parallelle lijnen in de hals of als het meerdere kinnen een zware set persoon zou naar voren vertonen door het kantelen van het hoofd. Is dit incompetentie of oneerlijkheid?
Afgezien van de schijnbare creationist neiging te misleiden door weglating, wat is hun verklaring voor dit patroon van embryonale overeenkomsten tussen gewervelde dieren die zich ontwikkelen tot heel andere volwassenen?
When it comes to shared characteristics between different groups of organisms creationists are fond of drawing (deeply flawed) analogies between living things and manufactured items like automobiles. They argue that the reason organism A and organism B share a characteristic is not due to common ancestry, but rather common design in the same way cars produced by two different car companies both have four wheels and pistons etc. Of course if we take that analogy seriously the question that immediately comes to mind is that if amniotes are like automobiles, then why do they begin life by developing an embryonic form that more resembles a submarine?
In reality their use of the term “common design” does nothing but relabel our ignorance. Worse it creates whole new sets of unanswerable questions. Designed by whom? Was it one designer or many (as in car companies)? How did he/she/it/they implement their design? Why did they design things the way they did? Why did they implement their designs in the timetable that they did? En maar door.
Het wordt nog erger als de “ontwerper” is transcendent en almachtig. Dit maakt het concept volledig toetsbaar (en dus onwetenschappelijk) als zodanig een schepper kan van alles maken, op welke wijze, om redenen die alleen bekend aan zichzelf. Het zou kunnen leiden tot een wereld bevolkt door evolutionair onmogelijke organismen zoals griffioenen en centauren , of het kan een wereld die lijkt precies alsof het vanzelfsprekend was ontwikkeld zonder goddelijke interventie (dat is wat blijkt het geval te zijn met onze wereld) te maken. Er is geen denkbaar observatie dat zou kunnen vervalsen “God zij heeft gedaan”.
Vergelijk dit met de theorie van gemeenschappelijke afstamming. Onder de evolutie van dit soort vreemd in elkaar geflanst, jerry-opgetuigd, structuren, waar de dingen worden gewijzigd uit de beschikbare materialen om heel andere functies dienen, zijn niet alleen verklaarbaar, maar op verzoek van de theorie.
Met andere woorden, als evolutie waar is, dan moeten we vinden dit soort dingen.
Nummer 3: De enige reden dat u belt ze kieuwspleten is omdat je veronderstellen evolutie.
Wat evolutionisten zou zien wat ze willen zien, dit argument is, vervalst door de historische record. Zoals eerder opgemerkt faryngeale spleten in amniote embryo’s werden “Kiemenspalten” (gill kloven) door hun ontdekker, Rathke, in tal van de 1820′s tientallen jaren voordat Charles Darwin bracht de evolutie in de wetenschappelijke mainstream. En tot 1859 de overwegend creationistische wetenschappers die onderzoek deed naar deze structuren gevolgd Rathke de voorsprong (* zie voetnoot) (zie ook mijn vriendDon Frack het commentaar hieronder:
Hoe verschillend de conformaties van de Vis, de Reptielen, het vogeltje en de warmbloedige viervoeter, kunnen aan de periode van hun looptijd, zijn ze nauwelijks van elkaar te onderscheiden in hun embryonaal stadium, en de vroege developement opbrengst enige tijd in op dezelfde manier Ze bezitten allemaal in eerste instantie de karakters van waterdieren;. en de Kikker houdt zelfs dit formulier in voor een aanzienlijke periode na het verlaten van het ei. De jonge kikkervisje is in waarheid een vis, of we de vorm en de acties van de instrumenten van de progressieve beweging, de regeling van zijn organen van de bloedsomloop en van de ademhaling, of de staat van de centrale organen van het zenuwstelsel te beschouwen.
[…] Birds, though destined to a very different sphere of action from either fishes or reptiles, are yet observed to pass, in the embryonic stage of their existence, through forms of transition, which successively resemble these inferior classes. The brain presents, in its earliest formation, a series of tubercles, placed longitudinally, like those of fishes, and only assuming its proper character at a later period. The respiratory organs are at first branchiæ, placed like those of fishes, in the neck, where there are also found branchial apertures similar to those of the lamprey and the shark ; and the heart and great vessels are constructed like those of the tadpole, with reference to a branchial circulation. In their conversion to the purposes of aerial respiration, they undergo a series of changes precisely analogous to those of the tadpole.
Mammalia, during the early periods of their developement, are subjected to all the transformations which have been now described; commencing with an organization corresponding to that of the aquatic tribes; exhibiting not only branchiæ, supported on branchial arches, but also branchial apertures in the neck ; and thence passing quickly to the conditions of structure adapted to a terrestrial existence. The developement of various parts of the system, more especially of the brain, the ear, the mouth, and the extremities, is carried still farther than in birds. Nor is the human embryo exempt from the same metamorphoses; possessing at one period branchiæ and branchial apertures similar to those of the cartilaginous fishes ,* a heart with a single set of cavities, and a brain consisting of a longitudinal series of tubercles; next losing its branchiæ, and acquiring lungs, while the circulation is yet single, and thus imitating the condition of the reptile; then acquiring a double circulation, but an incomplete diaphragm, like birds; afterwards, appearing like a quadruped, with a caudal prolongation of the sacrum, and an intermaxillary bone; and lastly, changing its structure to one adapted to the erect position, accompanied by a great expansion of the cerebral hemispheres, which extend backwards so as completely to cover the cerebellum. (Roget 1834, pp. 631-635,emphasis mine)
Please note that the above quote is from ) Animal and Vegetable Physiology Considered with Reference to Natural Theology , Vol. II of Treatise V of: The Bridgewater Treatise on the Power Wisdom and Goodness of God as Manifested in the Creation . As you might imagine, this was absolutely not a tome of atheistic evolution.
We zullen nu kort schetst de evolutie van de luchtwegen zoals in het embryo van de hogere gewervelde dieren. […] Rond de derde dag van de ontwikkeling van het kuiken, vier paar spleten of dwarse spleten waarneembaar zijn achter de mond, in de situatie of the branchial apertures of fishes ; and at the same time, the branchial vessels are developed from the aorta, as already described (§325). One of the apertures is intermediate between each pair of vascular arches just as in the gills of fishes and tadpoles. Nothing like branchial tufts, however, are developed; and the appearance described is very transitory , the vessels changing their direction and condition within two days. The development of perfect gills would have been useless, as the animal has not to maintain its own existence like the tadpole, but subsists, until the time of the perfect evolution of its respiratory system, upon the store of aliment furnished by the parent. It is evident, however, that the history of this evolution is so far the same as in Reptiles and Fishes. The lung first appears as a simple closed sac lying at the posterior and lowest part of the thorax; it soon becomes bifid, and presents a cavity, which does not, however, for some time communicate with the intestinal tube, the trachea and bronchi being last developed. The history of the evolution of these organs in the Mammalia is precisely analogous. It is usually at about the sixth of the period of uterine gestation that the rudiments of the branchial apparatus are seen, as marked by the shortness and thickness of the neck, the penetration of the sides of the pharynx by the branchial clefts , and the division of the aorta into vessels corresponding in number and distribution with the branchial arteries of fishes . These general features have been observed in the embryos of most orders of Mammalia, not excepting man himself; and they are probably common to all. A few days after the appearance of the fifth arch, which is the last developed, the neck begins to elongate, the apertures are closed gradually on the outside, while the vascular arches undergo those changes by which the permanent arterial branches arising from the heart are formed. The lungs in Mammalia are developed touch in the same manner as in Birds. They are not discernible before the period when the branchial apertures begin to close; a single mass is first perceived, which is soon divided into the rudiments of a right and left lung by a longitudinal groove; and the trachea and bronchi are subsequently developed as in birds. Scarcely a more beautiful illustration of the Unity of Design manifested in the creation of different classes of animals could be adduced than this hidden but not obscured correspondence … (Carpenter 1839, pp. 320-321,emphasis mine)
Please note that Carpenter’s repeated use of the term “evolution” here has nothing to do with evolutionary theory but rather is being used in the original sense of an “unfolding process”, as in embryological development. And of course as with Roget, Carpenter viewed these structures as an illustration of the “unity of design manifested in creation”. There is no reading evolution (in the modern sense) into the evidence there.
Perhaps the best example would be the zoologist/paleontologist Louis Agassiz (1807-1873) of Harvard University. Agassiz was a hard core (progressive) creationist who essentially believed that God specially created each species as they are (or were) throughout geologic time. However, despite his inveterate creationism he recognized the similarities between “fish” gills and the pharyngeal apparatus amniote embryos. In fact Agassiz, a creationist, is the only scientist I have ever read that actually claimed that the embryos of amniote have functional gills!
It may therefore be said with perfect propriety, that the higher Vertebrates undergo changes through which in different periods of their life, they resemble the lower ones; that there is a period when the young bird has the structure, not only the form, but the structure, and even the fins, which characterize the Fish. And of the young Mammals the same may be said. There is a period in the structure of the young Rabbit, (in which the investigations have been traced more extensively than in any other species,) when the young Rabbit resembles so closely the Fish, that it even has gills, living in a sac full of water breathing as Fishes do . (Agassiz 1849, p. 96, emphasis mine)
Again, Agassiz utterly rejected Darwin’s theory. For example at the conclusion of his review of Darwin’s On the Origin of Species he wrote that he considered the “transmutation theory” (evolution) to be “…a scientific mistake, untrue in its facts, unscientific in its method, and mischievous in its tendency.” (Agassiz 1860, p.154)
He never wavered from this position; nor did he apparently ever waver from his position regarding functional gills in amniote embryos:
I have myself watched carefully all the successive changes of development in the North American Axolotl, whose recently discovered metamorphoses have led to much discussion in connection with the modem doctrine of evolution. I can see no difference between this and other instances of metamorphosis. Certain organs, conspicuous in one phase of the animal’s life, are resorbed and disappear in a succeeding phase. But this does not differ at all from like processes in the toads and frogs, for instance; nor does it even differ essentially from like processes in the ordinary growth of all animals. The higher Vertebrates, including man himself, breathe through gill-like organs in the early part of their life. These gills disappear and give place to lungs only in a later phase of their existence. Metamorphoses have all the constancy and invariability of other modes of embryonic growth, and have never been known to lead to any transition of one species into another. (Agassiz 1874, p. 9, emphasis mine)
The careful reader will note that in this quote Agassiz is actually calling these structures “gill-like” in while in the course of denying that such things might be evidence for evolution!
Now, while Agassiz was wrong about “higher vertebrates” breathing through their “gills” (and is the only scientist I have ever seen make this claim), he clearly recognized the existence of these gill-like structures even though he was as far as one could be from being an evolutionist. This and the previous early 19th century quotes clearly illustrates the fact that recognition of these embryonic structures in amniotes as being gill-like, even to the point of referring to them as gill or branchial clefts, can be done independently of the acceptance of evolutionary biology. It is therefor does not follow that informally calling them “gill slits” is necessarily reading evolution into the evidence.
Amusingly while the creationist Agassiz had mistakenly claimed that amniote embryos breathe through their “gills”, the much maligned Ernst Haeckel had this to say on the subject:
In all Vertebrata already discussed [lampreys, sharks, bony fish] we found that they either always breathed through gills, or at least did so in early life, as in the case of Frogs and Salamanders. On the other hand, we never meet with a Reptile, Bird, or Mammal which at any period of its actual life breaths through gills , and the gill-arches and openings which do exist in the embryos are, during the course of their ontogeny, changed into entirely different structures, viz. into parts of the jaw-apparatus and the organ of hearing. (Haeckel 1902, p.302, emphasis mine)
So let’s look at the historical score card, with all the creationist caterwauling about how pharyngeal clefts never function as gills in amniotes and how calling them “gill slits” is an atheistic evolutionary deception hatched by Ernst Haeckel, in mind.
I. faryngeale spleten werden ontdekt en beschreven in amniote embryo’s bijna tien jaar voordat Ernst Haeckel werd zelfs geboren (1825 vs 1834).
II. Ze werden aangeduid als “gill kloven” door hun ontdekker, Rathke.
III. They were regularly called “gill clefts” or “branchial clefts” by creationist scientists in the 34 years between their discovery and the publication of Darwin’s On the Origin of Species (1859), demonstrating that referring to them in this manner is not simply a matter of reading evolution into the evidence.
IV. While creationist scientists commonly referred to the pharyngeal clefts of amniote embryos as “gill” or “branchial” clefts, they generally understood that they never function as gills.
V. There was one noted exception to number IV and it was arch creationist (pun intended) Louis Agassiz, not the evolutionist Ernst Haeckel (or any other evolutionist).
One, two, three, four, five strikes, you’re out!
As is so often the case the creationists apparent lack of curiosity about the natural world and ignorance about the history and philosophy of science, especially where it pertains to evolutionary theory, has led them up a blind alley.
As I’ve shown, by the time Darwin published On the Origin of Species , creationist biologists had for years been talking about these curious structures in amniote embryos that look a lot like the gills of fish and attempting to explain why they might be that way (“Unity of Design” etc.). The evidence from embryology was not something concocted after the fact by Darwin and his supporters to prop up an a priori belief in evolution (that’s how modern creationists work). Rather what Darwin did was take facts that were already known and gave them a logical, coherent explanation.
All Darwin essentially said was: “Hey you know those embryonic things you’ve been wondering about? I think I have a better explanation for why they exist.”
Just as the concept of the geologic column, and the pattern of fossil record upon which it is based, came to be part of the supporting evidence for evolution—even though its originators were creationists—so too did the recognized and established facts of embryology in the early 19 th century.
Conclusie
As with so many other things about the natural world, creationists are simply in denial about the evidence from comparative embryology. They have no coherent, testable, alternative explanation. And their “information” regarding the subject is a muddle of half-truths, misinformation and (willful) ignorance relayed in a fashion that begs the question as to whether incompetence or dishonesty is to blame.
Voetnoot
* A few people have pointed towards apparent misspellings of words in these 19th century quotes, however these are the spellings given in the originals and not the result (as is usually the case) of my lack of typing, English and/or proofreading skills.
Referenties:
Agassiz, Louis (1849) Twelve lectures on comparative embryology: delivered before the Lowell institute, in Boston, December and January, 1848-9 , Redding & Co.
Agassiz, Louis (1860) “Prof. Agassiz on the Origin of Species”, The American Journal of Science and Arts , 2nd Series, Vol. XXX, pp. 142-154
Agassiz, Louis (1874) Evolution and Permanence of Type, p. 9
Ascherson, Ferdinand Mauritius (1848) “ On Congenital Fistulæ of the Neck with a Succinct History of the Branchial Fissures in Mammals and in Birds ” in Clay, Charles (1849) The British Record of Obstetric Medicine and Surgery for 1849 , Vol. II
Butt, Kyle 2010 “ Shubin’s Subtly Deceptive “Gill Arches” “, Apologetics Press (website) downloaded on 5-29-2012
Carpenter, William Benjamin (1839) Principles of comparative physiology, Intended as an Introduction to the Study of Human Physiology and as a Guide to the Philosophical Pursuit of Natural History , John Churchill, Soho
Coyne, Jerry A. (2009) Why Evolution is True , Viking
Darwin, Charles (1860) in a Sept. 10 letter to Asa Gray, published in Darwin, Francis (editor) (1896) The Life and Letters of Charles Darwin , Vol. II, New York: D. Appleton and Company, p 131.
Gillis, Andrew J., et al. (2009) “ Shared developmental mechanisms pattern the vertebrate gill arch and paired fin skeletons “, Proceedings of the National Academy of Sciences , 106(14): 5720–5724
Grigg, Russel (1996) “ Ernst Haeckel: Evangelist for evolution and apostle of deceit “, Creation , 18(2):33-36
Haeckel, Ernst (1902) The History of Creation Vol. II (4th Edition), D. Appleton and Co.
Harrub, Brad (2001) “ Haeckel’s Hoax—CONTINUED! “, Apologetics Press (website) downloaded on 5-29-2012
Hunt, Paul et al. (1991) “The branchial Hox code and its implications for gene regulation, patterning of the nervous system and head evolution”, Development ; Supplement 2:63-77
Hunter, Cornelius (2010) “ Why Coyne is False “, Darwin’s God (Blog), downloaded on 5-28-2012
Kardong, Kenneth (2009) Vertebrates: Comparative anatomy, function, evolution (5th ed.), McGraw-Hill, New York, NY
Kuratani, Shigeru (2004) “ Evolution of the vertebrate jaw: comparative embryology and molecular developmental biology reveal the factors behind evolutionary novelty “, Journal of Anatomy 205(5): 335–347
MacDonald, RB et al. (2010) “Regulation of Dlx gene expression in the zebrafish pharyngeal arches: from conserved enhancer sequences to conserved activity”, Journal of Applied Ichthyology 26:187-191
Menton, David N. (1991) “ Human Embryo “Gill Slits” During Development “, bestbiblescience.org (web page), downloaded on 5-28-2012
Mitchell, Tommy & Mitchell, Elizabeth (2007) “ Something fishy about gill slits! “, Answers in Genesis ; website
Morris, John (1989) “ Does the Human Embryo Go through Animal Stages? ” Acts & Facts 18(8)
Okabe, Masataka & Graham, Anthony (2004) “ The origin of the parathyroid gland “, Proceedings of the National Academy of Science 101(51):17716-17719
Price, George McCready (1924) The Phantom of Organic Evolution , pp. 157-159
Prince, Victoria E. et al. (1998) “Zebrafish hox genes: expression in the hindbrain region of wild-type and
mutants of the segmentation gene, valentine”, Development 125:393-406
Rathke, Martin Heinrich (1825a) “Kiemen bei Säugetier,” Isis (1825), 747-749;
Rathke, Martin Heinrich (1825b) “Kiemen bei Vögeln,” Isis (1825), 1100-1101.
Roget, Peter Mark (1834) Animal and Vegetable Physiology Considered with Reference to Natural Theology , Vol. II (Treatise V of The Bridgewater Treatise on the Power Wisdom and Goodness of God as Manifested in the Creation), William Pickering, London
Schilling, T. (2003) “ Making Jaws “, Heredity , 90:3-5
Sherwin, Frank (2011) “ Design of Man: No Evolutionary Evidences “, Acts & Facts 40(1):16
Shubin, Neil (2009) Your Inner Fish: A journey into the 3.5 billion year history of the human body , Vintage Books, New York, NY
Stansfield, William D. (1977) The Science of Evolution , MacMillan Publishing Company, Inc., p.110
Takezaki, Naoko et al. (2004) “ The Phylogenetic Relationship of Tetrapod, Coelacanth, and Lungfish Revealed by the Sequences of Forty-Four Nuclear Genes “, Molecular Biology and Evolution 21(8):1512-1524
Tuttle, Albert H. (1884) “ The Relation of the External Meatus, Tympanum, and Eustachian Tube to the First Visceral Cleft “, Proceedings of the American Academy of Arts and Sciences , New Series Vol. XI, Whole Series Vol. xix, pp. 111-132 + plates
Venkatesh, Byrappa et al. (2001) “ Molecular synapomorphies resolve evolutionary relationships of extant jawed vertebrates “, Proceedings of the National Academy of Science 98(20):11382-11387
Wagner, Rudolph (1844) Elements of Physiology , Sherwood, Gilbert, & Piper, Paternoster Row, London (translated from the German by Robert Willis).
Wells, Jonathan (2000) Icons Of Evolution: Science Or Myth?: Why much of what we teach about evolution is wrong , Regnery Publishing Inc.
In “fish” the other arches support the gills while in amniote they become modified into other structures in the neck. For example the second arch develops into the hyoid bone in the throat and the stapes in the ear of mammals.
In “fish” the other arches support the gills while in amniote they become modified into other structures in the neck. For example the second arch develops into the hyoid bone in the throat and the stapes in the ear of mammals.


Figuur 1. Een reproductie van de tekeningen van Haeckel.
De gelijkenis van de embryo’s is gebaseerd op de vorm of morfologie van de organismen in de diverse stadia.
Al sinds enige tijd weet men dat er ook grote verschillen zijn in de ontwikkeling van ei naar volwassene. Het is daarmee altijd een vrij subjectieve waarneming gebleven. Het is wel duidelijk geworden dat er een stadium is tussen dat van het ei en de volwassen vorm waarin de organismen sterk op elkaar lijken. Dit wordt het fylotypische stadium genoemd en wordt verbeeld met behulp van een zandloper (zie figuur 2). De eerste fasen binnen het phylum verschillen erg van elkaar, maar naarmate men dichter bij een tussenliggend stadia komt lijken de organismen juist weer veel op elkaar. Bij het bereiken van de volwassen vorm verschillen ze weer.

fig2 & fig 3
Nu zijn er in 2010 twee studies voltooid die aantonen dat er gedurende de passage door de nauwe doorgang in de zandloper niet alleen morfologische overeenkomsten weerspiegeld worden, maar ook overeenkomsten in de expressie van de genen.
Een eerste studie met verschillende soorten fruitvliegjes toont aan dat de genen die tot uitdrukking komen tijdens de fylotypische fase minder uiteenlopen tussen de verschillende soorten. De genen die dan tot uitdrukking komen lijken meer op elkaar tussen de verschillende soorten. Het bleek ook dat de genen die belangrijk zijn voor de anatomische ontwikkeling de smalle doorgang van de zandloper volgen, terwijl genen die daar niet bij betrokken zijn niet de smalle doorgang nemen.
Een tweede studie op zebravisjes toont aan dat gedurende embryogenese het de oudste genen zijn die tijdens het fylotypische stadium worden uitgedrukt. Dit laatste gegeven toont dus aan dat de genen die tot uitdrukking komen tijdens de fasen waarin de organismen het meest op elkaar lijken de oudste zijn (figuur 3).
Het lijkt mij dat er sprake is van twee onderzoeken die hetzelfde resultaat opleveren. ‘Oude’ genen zijn genen die veel overeenkomsten met elkaar hebben en op basis van de overeenkomsten wordt een fylogenetische boom geconstrueerd. Gedurende de smalle doorgang van de zandloper bij fruitvliegjes zijn de genen die tot expressie komen de genen die het minst uiteenlopen. Dit betekent dus ook dat ze fylogenetisch dicht bij elkaar staan én het oudst zijn. De twee studies tonen vanuit verschillende invalshoeken hetzelfde mechanisme aan.
Hiermee is duidelijk dat Haeckel, hoewel hij de tekeningen wat overdreven had, toch iets ontdekt had. Blijkbaar is deze ‘smalle’ fase goed geconserveerd, wat betekent dat het een fase is die essentieel is voor de daaropvolgende stadia.
Uit: Nature 1, 2 Jerry Coyne‘s blog, PZMeyers, Panda’sThumb.
Haeckels biogenetische grondwet
Ernst Haeckel (hier al eens eerder voorgesteld) was 1 van de grote voorvechters van de evolutietheorie op het continent (Europa behalve de Britse eilanden) tijdens het leven van Charles Darwin.
Toch is deze man vooral de geschiedenis in gegaan als de promotor van een onjuiste wet; de biogenetische grondwet*, beter bekend onder de formulering: De ontogenese recapituleert de fylogenese.
Ontogenese is een moeilijk woord voor levensloop van een individu (van eicel tot bejaarde). Fylogenese is de (evolutionaire) levensloop van een soort (van 1cellige tot de mens, bijvoorbeeld).
Haeckel zag in het opgroeien van een organisme de geschiedenis van het leven op aarde zich herhalen. Ieder zoogdierembryo, bijvoorbeeld lijkt in een bepaald stadium op een vis. Haeckel zag de kieuwen zelfs zitten. Ook andere lichaamsdelen leken hun evolutionaire geschiedenis te herhalen.
Haeckel had het mis. Haeckel leek er vanuit te gaan dat er bij elke (kleine) stap in de evolutie een stukje aan de embryonale ontwikkeling werd toegevoegd. De oude voorgaande stadia (vis, amfibie, reptiel) bleven volgens Haeckel vrijwel intact en werden hoogstens wat sneller doorlopen. Wie beter kijkt ziet dat het niet zo gaat en dat Haeckel hier de zaak overdrijft.
Dat neemt niet weg dat in embryo’s van verwante soorten overeenkomsten te zien zijn die een aanwijzing kunnen geven over de loop van de afstamming. Aan de andere kant kunnen embryologen wellicht hun voordeel doen met de kennis van de evolutionaire geschiedenis bij het determineren van de anatomie van een embryo.
*Er zijn zelfs mensen(= meestal creationisten ) die beweren dat hij zijn anatomische tekeningen van embryo’s opgeleukt heeft (fraude!) om zijn theorie te ondersteunen.
Voor die mensen: kijk eens in een vogelboek waarin tekeningen van vogels staan zodat de vogelaar in staat is om de verschillende soorten te herkennen. Ik garandeer u dat u nergens deze vogels uit het boek zult vinden.
Haeckel liet de embryo’s gestileerd zien. Daartoe legde hij ze in vergelijkbare posen. Daarbij komt dat hij de voor het goede beeld van het embryo , storende dooiers en vliezen weg laat.
Haeckel’s ABC of evolution and development /
MICHAEL K. RICHARDSON and GERHARD KEUCK#
http://nsmserver2.fullerton.edu/departments/chemistry/Evolution_creation/Web/Richardson2002.pdf


Pingback: Extended Evolutionary Synthesis (EES) | Tsjok's blog